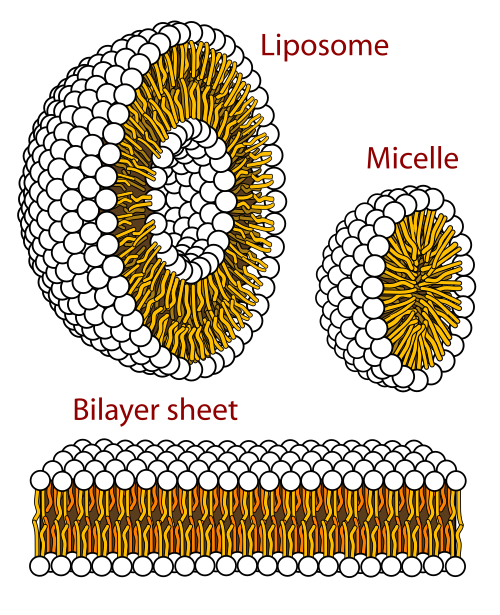
En biofísica y química coloidal , el polimorfismo es la capacidad de los lípidos para agregarse de diversas maneras, dando lugar a estructuras de diferentes formas, conocidas como " fases ". Esto puede manifestarse en forma de esferas de moléculas lipídicas ( micelas ), pares de capas enfrentadas ( fase lamelar , observada en sistemas biológicos como una bicapa lipídica ), una disposición tubular ( hexagonal ) o diversas fases cúbicas (Fd 3 m, Im 3 m, Ia 3 m, Pn 3 m y Pm 3 m son las descubiertas hasta ahora). También se han observado agregaciones más complejas, como las fases romboédrica , tetragonal y ortorrómbica .
Constituye una parte importante de la investigación académica actual en los campos de la biofísica de membranas (polimorfismo), la bioquímica (impacto biológico) y la química orgánica (síntesis).
La determinación de la topología de un sistema lipídico es posible mediante diversos métodos, siendo la difracción de rayos X el más fiable . Este método utiliza un haz de rayos X que, al ser dispersados por la muestra, generan un patrón de difracción en forma de anillos. La relación entre las distancias de estos anillos al punto central indica la(s) fase(s) presente(s).
La fase estructural de la agregación está influenciada por la proporción de lípidos presentes, la temperatura, la hidratación, la presión y la fuerza (y el tipo) iónico.
Fases hexagonales
En el polimorfismo lipídico, si la proporción de empaquetamiento de los lípidos es mayor o menor que uno, las membranas lipídicas pueden formar dos fases hexagonales separadas, o fases no lamelares, en las que se forman agregados tubulares largos según el entorno en el que se introduce el lípido.
Fase hexagonal I (H I )
Esta fase se ve favorecida en soluciones de detergente en agua y tiene una relación de empaquetamiento inferior a uno. La población micelar en una mezcla de detergente/agua no puede aumentar indefinidamente a medida que aumenta la proporción de detergente a agua. En presencia de bajas cantidades de agua, los lípidos que normalmente formarían micelas formarán agregados más grandes en forma de túbulos micelares para satisfacer los requisitos del efecto hidrofóbico. Estos agregados pueden considerarse micelas fusionadas. Estos túbulos tienen los grupos de cabeza polares orientados hacia el exterior y las cadenas de hidrocarburos hidrofóbicas hacia el interior. Esta fase solo se observa en condiciones únicas y especializadas, y probablemente no sea relevante para las membranas biológicas.
Fase hexagonal II (H II )
Las moléculas lipídicas en la fase HII se empaquetan de forma inversa al empaquetamiento observado en la fase hexagonal I descrita anteriormente. En solución, esta fase presenta los grupos polares de cabeza en el interior y las colas hidrofóbicas de hidrocarburos en el exterior. La relación de empaquetamiento para esta fase es mayor que uno, [ 1 ] lo que es sinónimo de un empaquetamiento de cono inverso.
Se formarán conjuntos extendidos de tubos largos (como en la fase hexagonal I), pero debido a la forma en que se empaquetan los grupos de cabezas polares, los tubos adoptan la forma de canales acuosos. Estos conjuntos pueden apilarse como tuberías. Esta forma de empaquetamiento puede dejar una superficie hidrofóbica finita en contacto con el agua en la parte exterior del conjunto. Sin embargo, este empaquetamiento, energéticamente favorable, aparentemente estabiliza esta fase en su conjunto. También es posible que una monocapa externa de lípidos recubra la superficie del conjunto de tubos para proteger la superficie hidrofóbica de la interacción con la fase acuosa.
Se sugiere que esta fase se forma a partir de lípidos en solución para compensar el efecto hidrofóbico. El empaquetamiento compacto de los grupos de cabeza de los lípidos reduce su contacto con la fase acuosa. Esto, a su vez, reduce la cantidad de moléculas de agua ordenadas pero no unidas. Entre los lípidos más comunes que forman esta fase se encuentra la fosfatidiletanolamina (PE), cuando posee cadenas de hidrocarburos insaturadas. El difosfatidilglicerol (DPG, también conocido como cardiolipina), en presencia de calcio, también es capaz de formar esta fase.
Técnicas de detección
Existen diversas técnicas para determinar qué fase está presente durante las perturbaciones realizadas en el lípido. Estas perturbaciones incluyen cambios de pH, cambios de temperatura, cambios de presión, cambios de volumen, etc.
La técnica más común para estudiar la presencia de fases fosfolipídicas es la resonancia magnética nuclear de fósforo (RMN de 31P). Mediante esta técnica, se observan patrones de difracción de polvo distintos y únicos para las fases lamelares, hexagonales e isotrópicas. Otras técnicas que se utilizan y que ofrecen evidencia definitiva de la existencia de fases lamelares y hexagonales incluyen la microscopía electrónica de criofractura, la difracción de rayos X , la calorimetría diferencial de barrido (DSC) y la resonancia magnética nuclear de deuterio (RMN de 2H).

Además, se ha demostrado que la microscopía electrónica de transmisión con tinción negativa es una herramienta útil para estudiar el comportamiento de fase de la bicapa lipídica y el polimorfismo en fase lamelar , micelar, liposoma unilamelar y estructuras lipídicas acuosas hexagonales , en dispersiones acuosas de lípidos de membrana . [ 2 ] Como la tinción negativa soluble en agua se excluye de la parte hidrofóbica (cadenas de acilo graso) de los agregados lipídicos, las porciones del grupo de cabeza hidrofílico de los agregados lipídicos se tiñen de oscuro y marcan claramente los contornos de los agregados lipídicos (véase la figura).
Véase también
Referencias
- ↑ Stuart, Marc y Boekema, Egbert. (2007). Dos mecanismos distintos de transición de vesícula a micela y de micela a vesícula están mediados por el parámetro de empaquetamiento de los sistemas fosfolípido-detergente, https://www.researchgate.net/publication/6124701_Two_distinct_mechanisms_of_vesicle-to-micelle_and_micelle-to-vesicle_transition_are_mediated_by_the_packing_parameter_of_phospholipid-detergent_systems#pf9
- ↑ YashRoy RC (1994) Desestabilización de la dispersión lamelar de los lípidos de la membrana tilacoide por la sacarosa. Biochimica et Biophysica Acta vol. 1212(1), pp. 129-133. https://www.researchgate.net/publication/15042978_Destabilisation_of_lamellar_dispersion_of_thylakoid_membrane_lipids_by_sucrose?ev=prf_pub
- JM Seddon, RH Templer. Polimorfismo de los sistemas lípido-agua , del Manual de Física Biológica, vol. 1, ed. R. Lipowsky y E. Sackmann. (c) 1995, Elsevier Science BV ISBN 0-444-81975-4
- Yeagle, P. (2005). La estructura de las membranas biológicas (2.ª ed.). Estados Unidos: CRC Press.
- Yeagle, P. (1993). Las membranas de las células (2ª ed.). Michigan: Academic Press.
- Gennis, RB (1989). Biomembranas: Estructura molecular y función. Michigan: Springer-Verlag.
- Biofísica
- Tensioactivos
- cristales líquidos
- Química coloidal